
分子生物学基础#
1. 微生物命名#
1.1. 分类#
分类
非细胞型:病毒、亚病毒
原核细胞型:细菌(支原体、衣原体、立克次氏体、放线菌和螺旋体)
真核细胞型:酵母、霉菌、原生动物、藻类
双名法
属名 + 种名(斜体)+ 命名者姓氏缩写(正体)
属名:首字母大写
种名:小写字母
名词所有格(表来源)
形容词主格(表性质)
1.2. 基因#
一般方法(三部分)
3 lower case letters (pathway)
1 capital letter (actual gene)
1 allele number
突变体
Superscripts |
Meaning |
Example |
Comment |
|---|---|---|---|
+ |
Wild type |
\(leuA⁺\)- |
Mutant |
ts |
temperature sensitive |
\(leuA^{ts}\) |
|
cs |
cold sensitive |
\(leuA^{cs}\) |
|
am |
amber mutation |
\(leuA_{am}\) |
|
um |
umber (opal) mutation |
\(leuA_{um}\) |
琥珀突变 |
oc |
ochre mutation |
\(leuA_{oc}\) |
赭石突变 |
R |
resistant |
\(rif^R\) |
|
Δ |
deletion |
\(ΔleuA\) |
|
- |
fusion |
\(leuA-lacZ\) |
|
: |
fusion |
\(leuA:lacZ\) |
|
:: |
insertion |
\(leuA::Tn10Ω\) |
|
Δ(gene)::(resistance) |
deletion/replacement |
ΔleuA::npt |
1.3. 表型#
first-letter capitalized
not italicized
DnaA⁻ the protein produced by the dnaA gene
LeuA⁻ the phenotype of a leuA mutant
2. 基因结构#
2.1. 基础概念#
基因(gene):即遗传因子,是产生一条多肽链或功能 RNA 所需的全部核苷酸序列,即带有遗传讯息的 DNA 片段;
顺反子(cistron):即结构基因,为决定一条多肽链合成的功能单位。顺反子在一定条件下与基因同义;
多顺反子 mRNA:含有编码一个以上蛋白质的编码信息,且,这些蛋白质均是以独立的多肽被翻译的 mRNA;
全酶(holoenzyme):一些酶除蛋白质外,还含有对热稳定的小分子物质。前者称为酶蛋白(apoenzyme),后者称为辅因子(cofactor)。只有酶蛋白和辅因子结合成完整的分子时,才具有活力;
辅酶(coenzyme):一类可将化学基团从一个酶转移到另一个酶上的有机小分子。与酶较为松散地结合,对于特定酶的活性发挥是必要的;
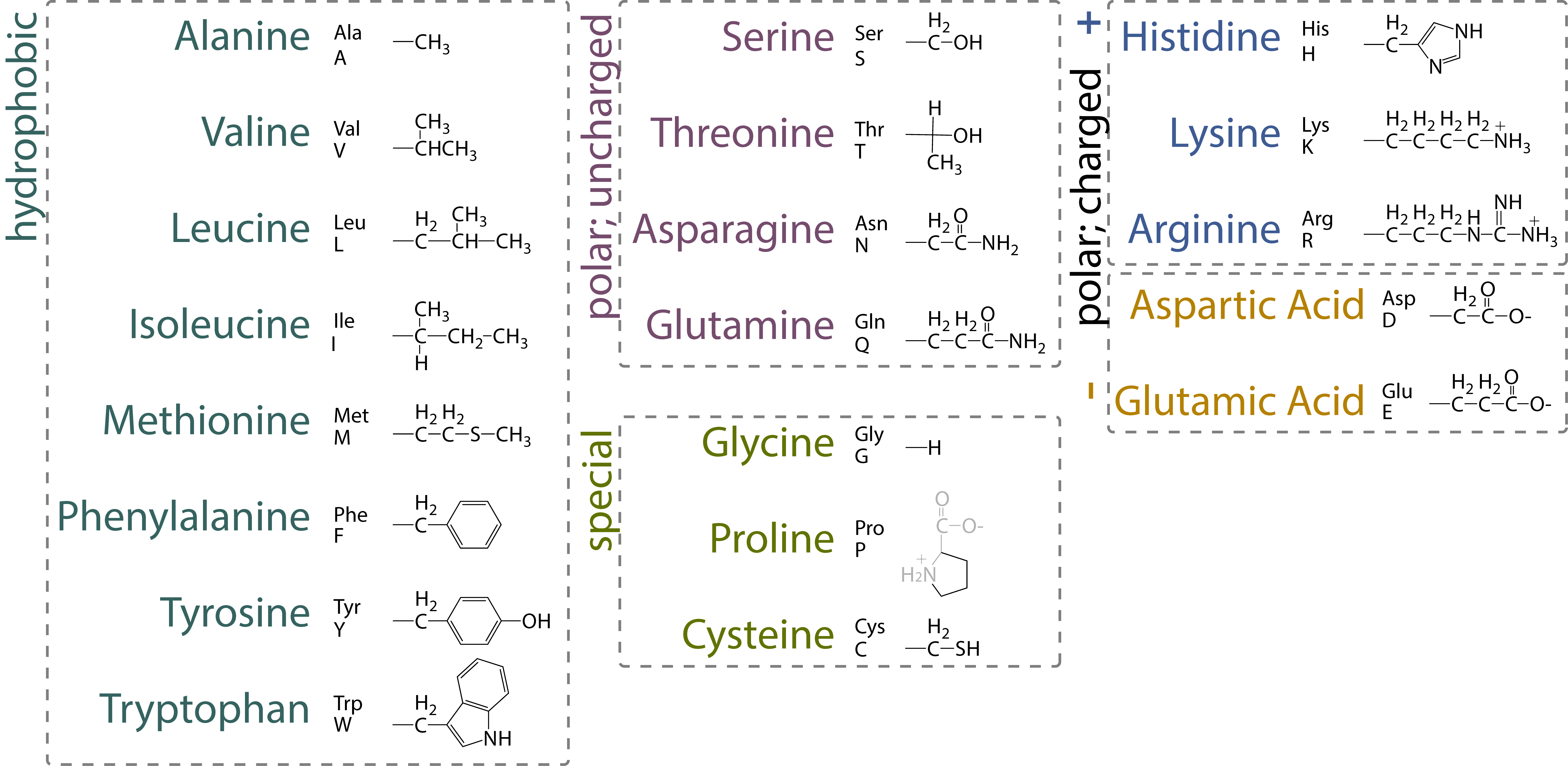
2.2. 复制、转录、翻译#
复制
以 DNA 为模板,在 DNA 聚合酶的催化作用下,将四种游离的 dNTP 按照碱基互补配对原则合成新链 DNA,为半保留复制,需要引物。
转录
以 DNA 为模版,在 DNA 指导的 RNA 聚合酶的作用下,将四种游离的 NTP 按照碱基互补配对的原则合成 RNA,为半不连续转录,需要引物。
翻译
以 mRNA 为模板,原料为 20 中游离的氨基酸,3 个碱基决定 1 个氨基酸。
2.3. ORF, CDS#
开放读码框(Open Reading Frame, ORF)是从一个起始密码子开始到一个终止密码子结束的一段序列;不是所有读码框都能被表达出蛋白产物或有生物学功能的蛋白;
CDS 必定是一个或多个 ORF;但,每个 ORF 不一定均是 CDS;
ORF = translation(理论可能)
CDS = transcription(事实存在);
2.4. 启动子(Promoter)#
位于编码区上游的非编码区。真核生物启动子包括下列几种不同顺序,能促进转录过程:
帽子位点
转录的起始位点;
TATA 框(Hogness box)
似于原核生物的 Pribnow 框,决定转录起点的选择,为 RNA 聚合酶的结合处;
一致顺序为 TATAATAAT;
约在基因转录起始点上游约 -30-50bp 处;
CAAT 框(CAAT box)
真核生物基因常有的调节区,控制转录的起始频率,可能亦为 RNA 聚合酶的结合处之一;
一致顺序为 GGGTCAATCT;
位于转录起始点上游约 -80-100bp 处;
GC 框(GC box)
转录因子 Sp1 能识别 GC 框且与之结合,其 N 端有激活转录的作用;
由 GGCGGG 组成;
位于 CAAT 框的两侧,有 2 个拷贝;
2.5. 增强子(Enhancer)#
又称远上游序列(far upstream sequence),一般都在 -100bp 以上;
对依赖于 TATA 框的转录和不依赖 TATA 框的转录都有增强效应,但对前者增强效应高;
增强子是通过启动子来增加转录的,有效的增强子可位于基因的 5’端,也可位于基因的 3’端,有的还可位于基因的内含子中;
增强子的效应很明显,一般能使基因转录频率增加 10-200 倍,有的甚至可高达上千倍;
2.6. 终止子(Terminator)#
在一个基因的末端往往有一段特定顺序,具有转录终止的功能,为反向重复序列,是 RNA 聚合酶停止工作的信号,反向重复序列转录后,可形成发夹式结构,且形成一串 U。发夹式结构阻碍了 RNA 聚合酶的移动,一串 U 的 U 与 DNA 模板中的 A 结合不稳定,从模板上脱落下来,转录终止。
3. 乳糖操纵子#
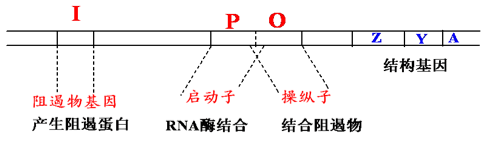
3.1. 操纵子(Operon)#
很多功能相关的结构基因串联排列在染色体上,由一个共同的控制区来操纵这些基因的表达,包含这些结构基因和控制区的整个核苷酸序列就称为操纵子。
1960∼1961 年,Monod 和 Jacob 首次提出”操纵子”学说(Lodish et al. 2000),获 1965 年诺贝尔生理学和医学奖;
1940 年,Monod 发现细菌在含葡萄糖和乳糖的培养基上生长时,细菌先利用葡萄糖,葡萄糖用完后,才利用乳糖;在糖源变换期,细菌的生长会出现停顿。即产生”二次生长曲线”。
3.2. 相关概念#
组成型表达(constitutive expression):在个体发育的任一阶段都能在大多数细胞中持续进行的基因表达,受环境因素的影响较少;
持家基因(housekeeping gene):表达产物通常是对生命过程必需的、必不可少的基因;
诱导(induction):在特定的环境信号刺激下,相应基因被激活,从而使基因的表达产物增加的过程;
阻遏(repression):在特定环境信号刺激下,相应基因被抑制,从而使基因的表达产物减少的过程;
协调表达(coordinate expression):在一定机制控制下,功能上相关的一组基因,无论其为何种表达方式,均需协调一致、共同表达,使各表达产物的分子比例适当,从而正常发挥功能的现象;
安慰诱导物(gratuitous inducer):能诱导酶的合成,但又不被分解的分子;
3.3. 组成#
三个特异性序列
操纵子 O(operator): 阻遏蛋白结合位点,指能被调控蛋白特异性结合的一段 DNA 序列;
启动子 P(promoter): 位于结构基因的上游,指能被 RNA 聚合酶识别、结合并启动基因转录的一段 DNA 序列;
CAP 结合位点:环 cAMP 受体蛋白(分解代谢物激活蛋白)结合位点;
一个调节基因
lac I:编码阻遏蛋白,能结合于操纵序列位点;
结构基因
结构基因(structural gene,SG):操纵元中被调控的编码蛋白质的基因
Z 编码 β-半乳糖苷酶:将乳糖水解成葡萄糖和半乳糖;
Y 编码 β-半乳糖苷透过酶:使外界的 β-半乳糖苷(如乳糖)能透过大肠杆菌细胞壁和原生质膜进入细胞内;
A 编码 β-半乳糖苷乙酰基转移酶:乙酰 CoA 上的乙酰基转到 β-半乳糖苷上,形成乙酰半乳糖;
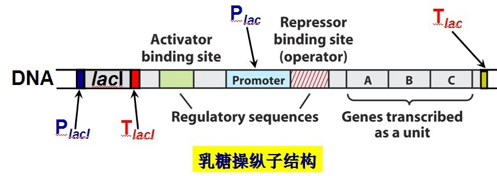
3.4. 多顺反子 mRNA#
多顺反子 mRNA 在细菌中很普遍。多顺反子 lac mRNA 中的 lacZ,lacY,lacA 经翻译生成的产物分别生成代谢分解乳糖的三种酶,且始终存在着一定的比例关系(Z:Y:A = 5:2:1)
lacZ,lacY,lacA 基因的转录是由 lacI 基因指令合成的阻遏蛋白 R 所控制;
lacI 一般和结构基因相毗连,但它本身具有自己的启动子和终止子,成为独立的转录单位;
由于 lacI 的产物是可溶性蛋白,按照理说是无需位于结构基因的附近,它是能够分散到各处或结合到分散的 DNA 位点上;
4. 乳糖操纵子的调控#
4.1. 阻遏蛋白的负调控#
无乳糖:lac 操纵子处于阻遏状态(repression);
有乳糖(presence of lactose):lac 操纵子即可被诱导;
诱导剂(inducer):别乳糖、半乳糖、IPTG(异丙基硫代半乳糖苷)
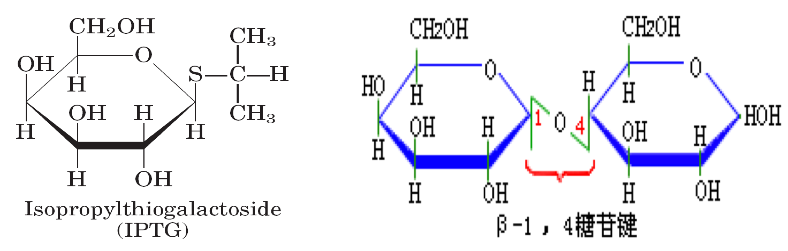
4.2. IPTG#
由于乳糖虽可诱导酶的合成,但又随之分解,产生很多复杂的动力学问题,因此人们常用安慰诱导物来进行各种实验,如异丙基-β-D-硫代半乳糖苷(IPTG);
IPTG 与自然的 β-半乳糖苷酶相似,且不能被酶分解,它的浓度在实验中不会改变。能在缺乏_lacY_ 基因下而有效地被运送。
半乳糖苷键中用硫代替了氧,失去了水解活性,但硫代半乳糖苷和同源的氧代化合物与酶位点的亲和力相同,IPTG 虽不为 β-半乳糖苷酶所识别,但它是 lac 基因簇十分有效的诱导物;
X-gal(5-溴 -4-氯 -3-吲哚-β-半乳糖苷)亦为一种人工化学合成的半乳糖苷,可被 β-半乳糖苷酶水解产生兰色化合物,因此可用作 β-半乳糖苷酶活性的指示剂;IPTG 和 X-gal 都被广泛应用在分子生物学和基因工程的工作中;
4.3. CAP 的正调控#
CAP(catabolite activator protein):分解代谢基因激活蛋白,又称为 CRP(cAMP receptor protein),lac 操纵子高水平转录必需的一个激活蛋白;
CAP(同二聚体),含 DNA 结合区 以二聚体的方式与特定的 DNA 序列结合,cAMP 结合区与 cAMP 特异结合,并发生空间构象的变化,形成 cAMP-CAP 复合物(有活性);
当 CAP 与 CAP 结合位点这段序列结合时,可激活 RNA 转录酶活性,使之提高 50 倍;
葡萄糖效应
当细菌在含有葡萄糖套和乳糖的培养基中生长时,通常优先利用葡萄糖。只有当葡萄糖消耗完,经过一段停滞期,在乳糖的诱导下半乳糖苷酶开始合成,细菌才能充分利用乳糖。葡萄糖的降解物能抑制腺苷酸环化酶活性,并活化磷酸二脂酶,因而降低 cAMP 的浓度。故葡萄糖存在时,cAMP 浓度低;仅在葡萄糖消耗完毕时,cAMP 浓度增高,CAP-cAMP 复合物形成(结合于 lac 操纵子 CAP 结合位点),才会促进转录。
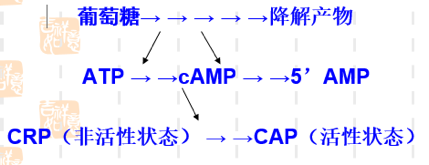
4.4. 协调调控#
由于 \(P_{lac}\) 是弱启动子,单纯因乳糖的存在发生去阻遏使 lac 操纵元转录开放,还不能使细菌很好利用乳糖,必需同时有 CAP 来加强转录活性,细菌才能合成足够的酶来利用乳糖;
关键条件:lac 操纵子的强诱导既需要有乳糖的存在又需要没有葡萄糖可供利用。
4.5. 阻遏蛋白和 CAP 的双重调控#
负调节与正调节协调合作
阻遏蛋白封闭转录时,CAP 不发挥作用;
如没有 CAP 加强转录,即使阻遏蛋白 R 从 P 上解聚仍无强大转录活性;
葡萄糖可降低 cAMP 浓度,阻碍其与 CAP 结合从而抑制转录;
本底表达
在细胞中透性酶等总是以最低量存在着,足以供给底物开始进入时的需要。即,操纵子有一个本底水平(basal level)的表达,即使没有诱导物的存在,也保持此表达水平(诱导水平的 1‰);有的诱导物是通过其他吸收系统进入细胞的。
5. 出芽酵母基因的命名#
Dominant gene: 3 upper case letters + 1 number
Recessive gene: 3 lower case letters + 1 number
Alleles: gene symbol + a hyphen + 1 italic Arabic number, e…g., act1-606, his2-1
Protein: Proteins are referred to by the relevant gene symbol, non-italic, initial letter uppercase and with the suffix p, e.g., Ade5p, Cdc28p
Phenotypes: a non-italic three-letter abbreviation corresponding to the gene symbol, initial letter uppercase. e.g., Arg⁻, Arg⁺.